|
Реферат: Зависимость интенсивности дыхания растительных продуктов от температурыРеферат: Зависимость интенсивности дыхания растительных продуктов от температурыМОСКОВСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ ПРИКЛАДНОЙ БИОТЕХНОЛОГИИ КАФЕДРА ХОЛОДИЛЬНОЙ ТЕХНИКИ И ТЕХНОЛОГИИ ТЕМА КУРСОВОЙ РАБОТЫ: ЗАВИСИМОСТЬ ИНТЕНСИВНОСТИ ДЫХАНИЯ РАСТИТЕЛЬНЫХ ПРОДУКТОВ ОТ ТЕМПЕРАТУРЫ ВЫПОЛНИЛ: Воробьев.А.Н ФАКУЛЬТЕТ:ХТиТ КУРС: 3 ГРУППА: 6 ПРИНЯЛА: Волдова.Т.А МОСКВА 2002 СОДЕРЖАНИЕ: 1.Введение. 2.Основные процессы и изменения, происходящие в продуктах растительного происхождения. 3.Процессы и изменения связанные с дыханием. 4.Хранение плодов в регулируемой газовой системе. 5.Охлаждение и хранение плодов и овощей в охлажденном состоянии. 6.Замораживание и хранение продуктов растительного происхождения в замороженном состоянии. 7.Режимы хранения. 8.Быстрое замораживание. 9.Замораживание клеток. 10.Количество вымерзающей воды. 11.Рекристаллизация. 12.Замораживание продуктов бестканевой структуры. 13.Водосодержание, теплоемкость и теплота замерзания некоторых продуктов. 14.Преимущества и недостатки. 15.Список используемой литературы. 1)Введение. В наше сложное время, с больной кризисной экономикой сохранение пищевых продуктов с наименьшими затратами и с большей эффективностью является одной из главных задач. Структура питания нуждается в улучшении технологии хранения пищевых продуктов, но улучшить структуру питания нельзя с помощью хладовой обработки, структуру питания можно лишь поддерживать с помощью хладовой обработки, что позволяет поднять качество продуктов растительного происхождения при хранении, и что в свою очередь позволяет увеличить сроки хранения продуктов, а так же стабильность хранения. При холодильной обработке и хранении в пищевых продуктах происходят сложные процессы и явления, приводящие к различным изменениям их структуры. Зависимостью интенсивности дыхания растительных продуктов от температуры, и исследованием влияния охлаждения (замораживания) на пищевые продукты я и решил заняться в данной курсовой работе. 2)Основные процессы и изменения, происходящие в продуктах растительного происхождения. Для понимания и совершенствования технологических процессов холодильного консервирования возникает настоятельная необходимость изучения изменений, происходящих в плодах и овощах при хранении, в зависимости от вида продукта, действия фитогормонов, устойчивости к фитопатогенным микроорганизмам и физиологическим заболеваниям. 3) ПРОЦЕССЫ И ИЗМЕНЕНИЯ, СВЯЗАННЫЕ С ДЫХАНИЕМ Условия холодильной обработки и хранения плодов и овощей должны быть таковы, чтобы понизить до минимума интенсивность дыхания, не нарушив при этом естественного течения метаболических процессов, поскольку только продукты растительного происхождения с нормальным дыханием обладают необходимой лежкоспособностью. Дыхание является основной формой диссимиляции — расщепления органических веществ. Это окислительный процесс, при котором потребляется кислород и выделяется углекислый газ. Обмен углекислого газа и кислорода происходит путем диффузии через устьица и частично через кутикулу. Главными дыхательными субстратами являются углеводы, жиры и белки. Около '/з количества углеводов в ткани расходуется при дыхании. Жиры и белки используются в меньшей степени, и их участие наиболее характерно для семян. Расщеплению макромолекулярных субстратов предшествует их гидролиз: поли- и дисахаридов до моносахаридов, жиров до глицерина и жирных кислот, белков до аминокислот. В результате реакции гликолиза из моносахаридов образуется пировиноградная кислота, затем вследствие окислительного декарбоксилирования пировиноградной кислоты — ацетил-КоА, который является также продуктом последовательной цепи превращений жирных кислот и аминокислот. В цикле Кребса ацетил-КоА подвергается реакциям декарбоксилирования и дегидрирования до полного расщепления. Образующиеся в цикле Кребса изолимонная, a-кетоглутаровая и яблочная кислоты являются непосредственными продуктами окисления. Процессы окисления весьма сложны и осуществляются через многоступенчатую систему специфических ферментативных реакций. Окисление может происходить непосредственным присоединением кислорода к дыхательному субстрату (А+0 → АО ); отщеплением водорода от субстрата (АН +Х → А+ХН ), при этом окисление субстрата АН и образование продукта окисления А происходят в результате восстановления вещества X; удалением электрона от заряженного иона (например, Fe —е → Fe ). В продуктах растительного происхождения происходят окислительные реакции всех трех типов, но процессы второго и третьего типов преобладают, так как они протекают без непосредственного участия кислорода. В основе современных представлений о механизме окислительных процессов в биологических объектах лежит теория Баха—Палладина, согласно которой дыхание клетки возможно лишь при наличии активированных форм кислорода и водорода, обладающих высокой реакционной способностью. Активация и передача водорода дыхательного субстрата происходят при участии ферментов дегидрогеназ, характеризующихся высокой специфичностью по отношению к окисляемому субстрату. По природе простетических групп дегидрогеназы делятся на пиридиновые и флавиновые. Коферментами пиридиновых дегидрогеназ являются НАД-никотинамидадениндинуклеотид и НАДФ-никотинамидаденин-динуклеотидфосфат, выполняющие функции переносчиков водорода. В основе действия пиридиновых дегидрогеназ лежит способность к обратимому гидрированию и дегидрированию пиридинового ядра, входящего в состав коферментов в виде амида никотиновой кислоты—витамина РР. Пиридиновые дегидрогеназы широко распространены в клетках продуктов растительного происхождения и являются в буквальном смысле универсальными окислительными системами. Флавиновые дегидрогеназы имеют коферменты, представляющие собой нуклеотидные производные рибофлавина — витамина В . Флавиновые ферменты весьма разнообразны, наиболее многочисленная их группа представлена дегидрогеназами, являющимися промежуточными переносчиками водорода в цепи окисления. Дегидрогеназы окисляют восстановленные пиридиновые основания и передают получаемые от них электроны по цепи в направлении к кислороду. Коферментами флавиновых дегидрогеназ, или, как их иногда называют, флавопротеидов (ФП), являются флавинмононуклеотид (ФМН) и флавинаде-ниндинуклеотид (ФАД). Последними в цепи окисления переносчиками водорода обычно выступают соединения, известные под общим названием коэнзимы Q. Этим наименованием обозначают ряд веществ, являющихся нейтральными липидами и представляющих собой соединение бензохинона с углеводородной боковой цепью, которая состоит из различного числа изопреновых групп. Известны коэнзимы Q с 10, 9, 8 и 7 изопреновыми группами. Будучи способными обратимо окисляться и восстанавливаться, коэнзимы Q являются связующим звеном в цепи переноса электронов от флавопротеидов к цитохромам. Активация кислорода в цепи окисления происходит под действием многочисленных ферментов (оксидаз), большинство из которых содержит в качестве простетических групп железопорфириновый комплекс, являясь Fe-протеидами. В цепи окисления из оксидаз основную роль играют цитохромы, состоящие из специфического белка и железопорфирина. Цитохромы способны обратимо окисляться и восстанавливаться благодаря ионам железа, входящего в их состав. Восстановление окисленных цитохромов происходит вследствие присоединения электронов KoQ либо флавопротеидов. Окисление цитохромов проходит через цепь, состоящую обычно из четырех различных цитохромов. Последним в цепи является цитохром Аз, называемый цитохромоксидазой, окисление которого происходит непосредственно кислородом воздуха. Как известно, способность какого-либо соединения присоединять или отдавать электроны определяется в основном соотношением величин окислительных потенциалов данного соединения и соединения, с которым оно взаимодействует. Причем реакция проходит тем энергичнее, чем меньше различие между потенциалами взаимодействующих веществ. К окислительному потенциалу дыхательного субстрата наиболее близки потенциалы, свойственные пиридиннуклеотидам, а к потенциалу кислорода, имеющего наибольшую величину,— потенциал цитохромоксидазы. Исходя из этого, считают, что первый этап окисления субстратов осуществляется с участием пиридиновых дегидрогеназ, второй — флавиновых дегидрогеназ, третий — коэнзима Q. Причем на этих этапах происходит перенос двух атомов водорода. Далее имеет место перенос непосредственно электронов через систему цитохромов на кислород. Процесс окисления субстрата в дыхательной цепи сопровождается фосфорилированием АДФ и запасанием энергии в форме АТФ. В цепи окисления есть три участка фосфорилирования: участок окисления флавопротеидов, участок перехода электронов от цитохрома В к цитохрому С и участок перехода электронов от цитохрома А к цитохрому Аз (рис. 1).
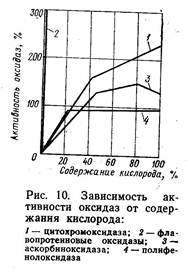
фосфорилированием и экспериментальное доказательство этого были даны академиком В. А. Энгельгартом. Как правило, процессы дыхания происходят в митохондриях клеток. Каждая стадия дыхания осуществляется мультиферментной системой, закрепленной на внутренней митохондриальной мембране. Около 1/4 белков внутренней мембраны являются ферментами, участвующими в переносе электронов и окислительном фосфорилировании. К ним относятся флавопротеиды, цитохромы, ферменты, участвующие в образовании АТФ. Флавопротеиды и цитохромы располагаются в мембране в виде отдельных групп, каждая из которых содержит все необходимые ферменты и функционирует самостоятельно. Такие группы, называемые дыхательными ансамблями, равномерно распределены по всей плоскости мембраны. Они весьма чувствительны к действию различных внешних воздействий. Под влиянием низких температур, ядов, ионизирууэщей радиации нарушается целостность митохондриальных мембран, и в этом случае сопряжение дыхания утрачивается, или происходит разобщение дыхания. Процессы фосфорилирования более чувствительны. Поэтому при разобщении дыхания окислительные процессы часто не нарушаются, но при этом энергия окисления не превращается в энергию АТФ, а рассеивается в форме тепла. Разобщение дыхания представляет собой необратимый процесс, являющийся признаком гибели клетки. Однако растительные ткани вследствие наличия у них специальных приспособительных механизмов способны переносить неблагоприятные внешние условия. Дыхательные процессы, протекающие в клетках, обладают рядом особенностей, обусловленных тем, что продукты растительного происхождения лишены стабильной внутренней среды, например парциального давления кислорода и углекислого газа, температуры, и изменяются под действием непостоянных и крайне изменчивых факторов внешней среды. Б. А. Рубин считает, что для структуры аппарата растений характерны три принципиальные особенности: 1. Принцип множественности (мультипринцип) при построении окислительных систем, обусловленный тем, что в клетке имеется, как правило, не один, а несколько ферментов, выполняющих одну и ту же функцию. 2. Полифункциональность каталитических систем, или наличие ферментов, обладающих не одним, а несколькими свойствами. 3. Рассредоточенность (делокализованность) аппарата. Вследствие этих особенностей процессы окисления биологических объектов могут осуществляться различными путями, или, как говорят, для растительных тканей характерно наличие альтернативных механизмов окислительного обмена, что играет исключительную роль как фактор адаптации, обусловливающий способность тканей сохраняться и нормально функционировать в условиях изменчивой внешней среды, а также при действии различных неблагоприятных факторов. Схема окисления дыхательного субстрата в клетках разных тканей различна и зависит от их физиологического состояния и условий существования. Так, в отдельные периоды жизненного цикла плодов, например при созревании, клетки нуждаются в дополнительном притоке энергии. При этом в качестве дополнительного дыхательного субстрата используется янтарная кислота, окисление которой происходит более коротким путем, минуя НАД, непосредственно через ФП на КоQ. При этом сукцинатдегидрогеназа, относящаяся к флавиновым ферментам, строго специфична по отношению к янтарной кислоте. Обладая способностью акцептировать водород непосредственно от субстрата и передавать его на КоQ, сукцинатдегидрогеназа образует боковую транспортную цепь окисления. Кроме того, в клетках имеются альтернативные окислительные механизмы, образующие сопряженные цепи с флавопротеиновыми ферментами и цитохромами. Среди них главную роль играют оксидазы, содержащие в молекуле ионы железа или меди. В группу Fe-протеидов наряду с флавопротеиновыми ферментами и цитохромами входят каталаза и пероксидаза. В прохождении дополнительного, альтернативного, пути окисления особенно велика роль пероксидаз, разлагающих перекись водорода с освобождением активного атомарного кислорода. Пероксидазы, встречающиеся в клетках продуктов растительного происхождения, весьма многочисленны и окисляют различные вещества, в том числе практически все фенолы, ароматические амины и аскорбиновую кислоту. Их активность возрастает при изменении условий внешней среды. Так, в паренхимной ткани плодов при пониженных температурах хранения или недостаточной обеспеченности кислородом происходит возрастание активности пероксидазы. К группе Сu-протеидов относятся различные полифеноло-ксидазы и аскорбиноксидаза. В основе их действия лежит обратимое окисление одновалентной меди в двухвалентную. Полифенолоксидазы представляют собой ферменты, окисляющие в присутствии молекулярного кислорода различные фенолы и их производные с образованием соответствующих хинонов. Полифенолоксидазы катализируют прямое окисление субстратов атмосферным кислородом и участвуют в различных окислительных процессах, в том числе в окислении аминокислот, полипептидов, белков и аскорбиновой кислоты. Активность полифенолоксидаз возрастает при механическом повреждении плодов, воздействии низких температур, кристаллизации воды при замораживании и т. д. Потемнение мякоти многих плодов (яблок, груш, абрикосов и др.), а также клубней картофеля, наблюдаемое при разрезании, очистке и механическом повреждении, происходит вследствие возрастания активности полифенолоксидаз и нарушения координации реакций окисления и восстановления, в результате чего в ткани накапливаются стойкоокрашенные продукты. Аскорбиноксидаза обладает строгой специфичностью по отношению к субстрату и окисляет l-аскорбиновую кислоту, превращая ее в дегидроформу. Являясь переносчиком водорода, аскорбиновая кислота тесно связана со всей системой ферментов, участвующих в дыхательных процессах. Полифенолоксидазы и аскорбиноксидаза принимают участие в окислительных процессах на заключительном этапе дыхания, заменяя цитохромы. На рис. 2 показаны возможные альтернативные пути процесса окисления, которые в. растительных клетках происходят различным образом в зависимости от видовых особенностей ткани, физиологического состояния данного органа, условий внешней среды и других факторов. Различная активность оксидаз в условиях измененной газовой среды представлена на рис. 3. Наибольшее сродство к кислороду имеет цитохромоксидаза, ее действие проявляется при содержании кислорода в атмосфере всего 1 %. Глубинные слои паренхимной ткани плодов и овощей, для которых характерно пониженное содержание кислорода, более богаты цитохромоксидазой, нежели поверхностные.
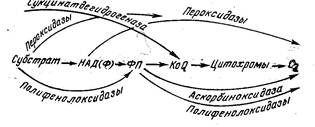
Рис. 2. Альтернативные пути окисления Ферменты, участвующие в альтернативных процессах
окисления, локализованы в различных структурных элементах клетки. Пероксидазы,
полифенолоксидазы, аскорбиноксидаза сосредоточены главным образом в
цитоплазме; вся система ферментов, обеспечивающая окислительное
фосфорилирование, обнаружена в ядрах и хлоропластах. Благодаря такой
делокализации ферментов дыхательной цепи практически все структурные составляющие
клетки могут быть источниками того или иного количества энергии и
использоваться ею в зависимости от условий внешней среды. О характере и интенсивности дыхания судят по количеству выделяемого клетками углекислого газа либо по величине дыхательного коэффициента ДК: ДК=Vсо,/Vо„ где Vco, Vo - объемы выделившегося углекислого газа и поглощенного кислорода. Величина дыхательного коэффициента определяется прежде всего химической природой субстрата. При окислении углеводов ДК=1 в соответствии с суммарным уравнением окисления глюкозы С Н О + 60 -> 6СО + 6Н О + Q. Число выделившихся грамм-молекул углекислого газа равно числу поглощенных грамм-молекул кислорода, а согласно закону Авогадро грамм-молекулы всех газов занимают одинаковый объем. При дыхании число выделившихся молекул углекислого газа равно числу атомов углерода в молекуле субстрата. Поэтому число использованных молекул кислорода на атом углерода в субстрате возрастает с увеличением числа Н-атомов и снижается с увеличением содержания O-атомов в молекуле субстрата. Если субстратами дыхания являются высшие карбоновые кислоты или аминокислоты, относительно богатые водородом и бедные кислородом, то ДK<1. Для жиров ДК~0,7, для белков ДК~0,8. В качестве примера приведем окисление стеариновой кислоты: С Н О + 26O -»18СО + 18Н О; ДК = 18/26 = 0,69. Если дыхание происходит с использованием низших ди- и трикарбоновых кислот (яблочной, лимонной, винной и щавелевой), богатых кислородом, то ДК>1. Так, при окислении яблочной кислоты С Н О + ЗО -» 4СО + ЗН О; ДК. = 4/3 = 1,33. Интенсивность дыхания и соответственно величина дыхательного коэффициента молодых развивающихся тканей, например меристематических, тканей прорастающих семян выше, чем зрелых плодов. У тканей одного и того же плода или овоща интенсивность дыхания периферических тканей, как правило, выше. Ниже приведена интенсивность дыхания [в мг СO2/(г*сут)] различных тканей при температуре 15—20 °С.
Лимон целые плоды кожура мякоть Картофель целые клубни кожица покоящиеся глазки мякоть При понижении температуры интенсивность дыхания плодов снижается и соответственно дыхательный коэффициент уменьшается. Так, по данным Ф. В. Церевитинова, дыхательный коэффициент для яблок сорта Канада составляет 1,14 при 30 °С и 0,88 при 0 °С. У большинства продуктов растительного происхождения дыхание прекращается при температуре около —10 °С. У растений, переносящих в естественных условиях суровые зимы, дыхание отмечается при —20°С и более низких температурах. Дыхание зависит от газового состава окружающей атмосферы. При понижении концентрации кислорода интенсивность дыхания снижается, при увеличении возрастает. Интенсивность дыхания снижается вследствие увеличения концентрации углекислого газа. Когда содержание кислорода в окружающей атмосфере уменьшается ниже определенного предела или при полном отсутствии кислорода, клетки продуктов растительного происхождения переходят к так называемому интрамолекулярному или бескислородному дыханию, представляющему собой в химическом отношении брожение. Брожение наряду с дыханием является основной формой диссимиляции веществ в организме. При дыхании суммарный выход энергии велик, а конечными продуктами являются неорганические соединения углекислого газа и воды. При брожении выход энергии значительно меньше, органический материал не распадается до конца и накапливаются относительно богатые энергией органические вещества. Для большинства тканей продуктов растительного происхождения характерна способность к спиртовому брожению, для некоторых — к молочнокислому. Начальные стадии дыхания и брожения одинаковы, но в анаэробных условиях процесс диссимиляции углеводов прекращается на стадии гликолиза при молочнокислом брожении либо на стадии образования ацетальдегида и этанола при спиртовом. Переход к интрамолекулярному дыханию у растений может быть частичным, что определяется не только составом газовой среды, но и физиологическим состоянием организма. Так, бескислородное дыхание отмечается в период созревания плодов. При полном отсутствии кислорода переход исключительно к интрамолекулярному дыханию приводит к гибели организма, так как накапливающийся этанол губительно действует на живую клетку. 4)ХРАНЕНИЕ ПЛОДОВ В РЕГУЛИРУЕМОЙ ГАЗОВОЙ СРЕДЕПри хранении плодов практически единственной формой их взаимодействия с окружающей средой является дыхательный газообмен. Поэтому их жизнедеятельность в роцессе хранения в значительной степени определяется составом окружающей атмосферы. Изменяя определенным образом газовую среду, например повышая содержание углекислого газа и уменьшая содержание кислорода, можно снизить интенсивность обменных процессов в хранимых плодах, для того чтобы отделить состояние климактерия и старения плода. На подавлении жизнедеятельности путем создания определенного состава атмосферы основано хранение плодов в регулируемой газовой среде. Для паренхимной ткани плодов характерно наличие обширных межклеточных пространств, заполненных газом. Газовый состав внутренней атмосферы плодов устанавливается в результате динамического равновесия между интенсивностью потребления кислорода и выделением углекислого газа, с одной стороны, и скоростью диффузии этих газов — с другой. Обмен с окружающей средой происходит через кутикулу, которая затрудняет циркуляцию газов, вследствие чего внутри плода создается своеобразная газовая атмосфера с повышенным содержанием углекислого газа и пониженной концентрацией кислорода. Характерно, что состав атмосферы в межклетниках неодинаков: в расположенных ближе к поверхности межклетниках состав атмосферы более близок к составу окружающей среды. Газовый состав в межклетниках у разных плодов различен, изменяется с их возрастом и зависит от температуры и наружной атмосферы. Кутикула плодов разных сортов различается по толщине и составу, в результате чего поступление внутрь кислорода и его концентрация в межклетниках и клеточном соке также варьируются. В связи с этим одни плоды будут испытывать недостаток кислорода или избыток углекислого газа, тогда как для других эта же газовая среда окажется благоприятной. Кислородные оптимумы дыхания неодинаковы на разных этапах развития плода, а также смещаются под воздействием температурного фактора. При 30 °С оптимальной является 5 %-ная концентрация кислорода, при температуре 10 °С и ниже—1 %-ная. Повышенные концентрации углекислого газа особенно благоприятны для задержки обменных процессов в плодах в постклимактерическом состоянии. |
|
||||||||||||||||||||||||||||||||
|
|||||
Рефераты бесплатно, реферат бесплатно, рефераты на тему, сочинения, курсовые работы, реферат, доклады, рефераты, рефераты скачать, курсовые, дипломы, научные работы и многое другое. |
||
При использовании материалов - ссылка на сайт обязательна. |
||